Post 2: The Geologic Column
When the geologic column was first being mapped out by geologists they could only establish relative dates of the position of formation of a given layer in the column based on the premise that 'layers buried further down' = 'older than newly formed surface layers'… with care being taken to ensure you weren’t analyzing something like an overthrust where one section of plate has pushed up on top of another one. Then came radiometric dating which allowed them not only to independently test that hypothesis but to assign specific age values to each of those layers… resulting in the modern understanding of the geologic column. For example, in Glenn Morton’s article on the geologic column at Talk.Origins (
http://www.talkorigins.org/faqs/geocolumn/) one of the references used is a well dug in North Dakota to a depth of over 15 thousand feet. The following layers were encountered at the respective depths: (Fm = Formation, Lm = Limestone, Grp = Group)
Tertiary Ft. Union Fm ...............................100 feet
Cretaceous Greenhorn Fm .......................4910 feet
Cretaceous Mowry Fm............................ 5370 feet
Cretaceous Inyan Kara Fm.......................5790 feet
Jurassic Rierdon Fm................................6690 feet
Triassic Spearfish Fm..............................7325 feet
Permian Opeche Fm................................7740 feet
Pennsylvanian Amsden Fm.......................7990 feet
Pennsylvanian Tyler Fm...........................8245 feet
Mississippian Otter Fm.............................8440 feet
Mississippian Kibbey Lm...........................8780 feet
Mississippian Charles Fm..........................8945 feet
Mississippian Mission Canyon Fm................9775 feet
Mississippian Lodgepole Fm.....................10255 feet
Devonian Bakken Fm.............................11085 feet
Devonian Birdbear Fm............................11340 feet
Devonian Duperow Fm...........................11422 feet
Devonian Souris River Fm.......................11832 feet
Devonian Dawson Bay Fm.......................12089 feet
Devonian Prairie Fm...............................12180 feet
Devonian Winnipegosis Grp.....................12310 feet
Silurian Interlake Fm..............................12539 feet
Ordovician Stonewall Fm........................13250 feet
Ordovician Red River Dolomite.................13630 feet
Ordovician Winnipeg Grp........................14210 feet
Ordovician Black Island Fm.....................14355 feet
Cambrian Deadwood Fm.........................14445 feet
Precambrian.........................................14945 feet
The article also includes 25 other sites where the entire column has been observed.
The span of ages since associated with each of those eras since the advent of radiometric dating are:
Tertiary –------------------ 1.8 million -> 65 million years old
Cretaceous --------------– 65 million -> 145 million years old
Jurassic ----------------– 145 million -> 205 million years old
Triassic ----------------–205 million -> 250 million years old
Permian –---------------- 250 million -> 290 million years old
Pennsylvanian –---------- 290 million -> 325 million years old
Mississippian –------------ 325 million to 355 million years old
Devonian –--------------- 355 million -> 420 million years old
Silurian –----------------- 420 million -> 445 million years old
Ordovician –-------------- 445 million -> 490 million years old
Cambrian Deadwood Fm –- 490 million -> 545 million years old
Precambrian –------------------------ 545+ million years old
So, the further down we go, the older the dates we see. Exactly as predicted. But that isn’t the only indicator to consider, there is also the fossil composition of the geologic column. I’ll do a quick overview of which fossils are found in which layers for now... starting with what are dated as the oldest layers and progressing through to the youngest. Note that the precise locations of many of these "earliest known fossil" finds are constantly being adjusted to some degree as more and more fossil finds come in and the body of what is known is added to.... for example, not too many years ago the earliest known multicellular fossils were early Cambrian (540 million years old) but then someone found some in layers about 20 million years older than that and the date of the earliest known multicellular fossils got shifted back a few percent into the late Precambrian. This is to be expected... and will certainly continue to happen in the future.
Precambrian
--In the oldest dated layers of rock in the Precambrian there has never been a fossil found. Of anything. Ever.
--As we move to newer layers in the Precambrian we start finding fossils of single celled organisms at about the 3.5 billion year mark. They appear to be prokaryotes. We find fossils of
nothing else.
--In still newer layers we begin finding fossils of what seem to be eukaryotic single celled organisms. (Prokaryotes have cell structures that lack mitochondria and nuclei, eukaryotes incorporate mitochondria and nuclei).
--In the late Precambrian layers leading up to the Cambrian, we begin finding fossils of small, simple, multicellular organisms (for example: the Ediacaran fauna) and also fossils of what appear to be simple chloroplasts.
Cambrian
--Once we reach the Cambrian we have the “Cambrian Explosion”. Keep in mind that this “explosion” takes tens of millions of years… some people have the unfortunate tendency to think this means that: *poof*… a bunch of different animals just all showed up simultaneously.
--By the end of the Cambrian we see the emergence of the earliest representatives of most existent phyla. Note that for the most part they look absolutely nothing like modern representatives of those phyla… another point on which people have an unfortunate tendency to become confused. They think that (for example) because we have brachiopods today, and brachiopods showed up in the Cambrian, therefore modern brachiopods have been around since the Cambrian. This is just plain wrong.
--Among the organisms first appearing in the Cambrian are: Arthropods (trilobites!), Molluscs, Chordates (near the end of the Cambrian), Brachiopods, etc…
Ordovician
--The first fossil Bryozoans show up in the Ordovician, little colonies of interconnected aquatic organisms that ten to inhabit rock surfaces, etc…
--The first coral fossils.
--Earliest jawless fish, although there is some evidence they may have shown up in the late Cambrian.
Silurian
--Fossils of jawless fish are abundant and diverse. Earliest fossils of fish with jaws are found.
--The first fossil evidence of
any land organisms. Fungi, and also
cooksonia, the earliest known plant with a vascular network.
--By the late Silurian we also find primitive fossil arachnids and centipedes.
--Note that we have four and a half billion years of rock layers and find no evidence of anything non-aquatic until the latest ten percent of them.
Devonian
--Earliest fossils of tetrapods (amphibians).
--Towards the end of the Devonian we find the earliest fossils of seed bearing plants.
Mississippian and Pennsylvanian (Combined = Carboniferous)
--Earliest amniote fossils.
--Tetrapod fossils become increasingly diverse.
--Land based plant fossils also diversify.
--Towards the end of the Pennsylvanian the earliest diapsid fossils are found (animals with two fenestrae. Ie: reptiles)
Permian
--For most of the Permian increasingly diverse examples of the previously mentioned groups are found…
--At the end of the Permian there appears to be a large scale mass extinction event. A massive number of species found in the fossil record prior to this time cease to be found at any point later (goodbye trilobites… you had a good run…).
Triassic
--The emergence of the dinosaurs in the fossil record. The popular giant versions are not found at this point in the fossil record… Triassic dinosaur fossils consist of smaller representatives of that group.
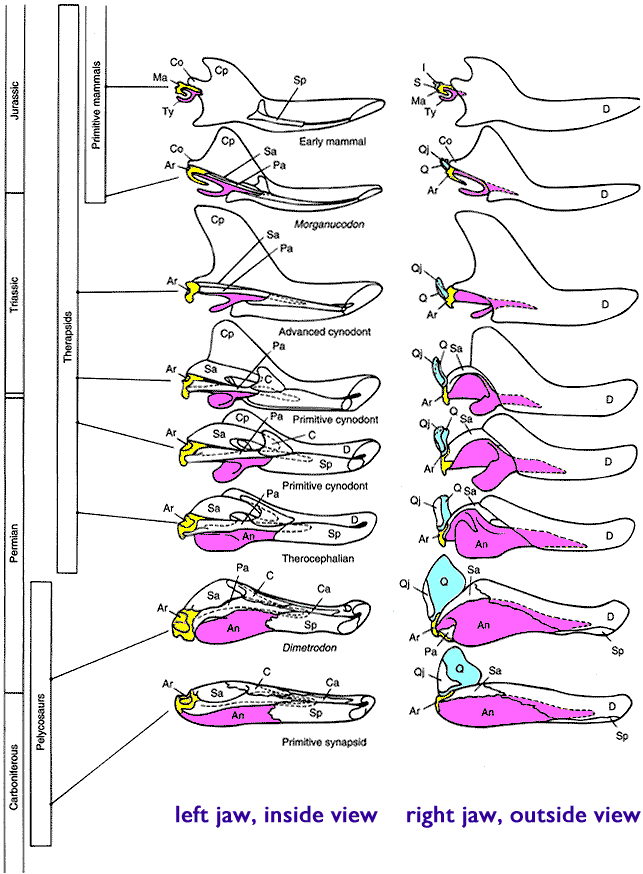
--Towards the very end of the Triassic we find the first fossils of small mammals.
Jurassic
--Dinosaur fossils get bigger and more diverse.
--Crocodiles show up.
--By this point in the fossil record aquatic life is extremely diversified. Sharks, rays, fish, squid, ammonites, all kinds of aquatic plants…
--Land plants also become increasingly diverse, as well as mammals.
--Near the end of the Jurassic, we have Archeopteryx. Clearly reptilian… but feathered.
Cretaceous
--Fossils of flowering plants (angiosperms) appear.
--Fossils of modern looking versions of some mammals and insects.
--Dinosaur fossils continue to diversify. A crowd favorite, T-rex, makes it’s appearance in the cretaceous layers. Unfortunately for it:
--At the very end of the Cretaceous there is another apparent mass extinction event, the K-T event. No further fossil evidence of dinosaurs and many other species found previous to this point in the fossil record are found in later layers. The K-T boundary marks the end of the Cretaceous period in the geologic column, a thin layer in the column with heavy iridium concentrations, found worldwide, leading to the hypothesis that there was a massive meteor/asteroid strike at this time which kicked up enough impact debris to lay down a coating over the entire surface of the planet. This event is more well known than the Permian mass extinction, even though it appears to have wiped out a smaller percentage of the extant species than did the Permian event.
Tertiary
--Within the Tertiary layers we find fossils of modern animal forms. Modern angiosperm plants, mammals, ray-finned fish, birds, etc…
--We begin finding the first primate fossils right near the KT boundary. They’re small… in appearance they resembled something like a squirrel. The first prosimian fossils (for example:
Smilodectes) show up in the early Tertiary layers. The first ape and monkey fossils begin appearing in the mid-Tertiary layers. (
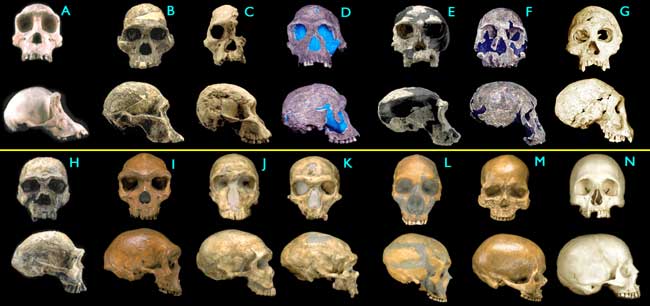
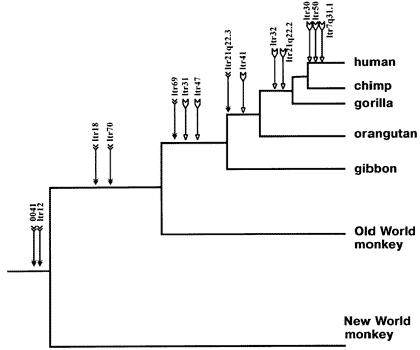
Apidium, Aegyptopithecus, etc…). Ape and monkey fossils continue to diversify throughout the later layers of the Tertiary. Approaching the end of the Tertiary the first Hominid fossils are found, dating back approximately 5-6 million years. It bears thinking on that hominid fossils occupy only the upper approximate 1% of the geologic column. Fossils of homo sapiens are not found in Tertiary layers.
Note that the well mentioned earlier was dug in a basin, beginning below the very upper layers (the Quaternary layers which are dated at 1.8 million years to present) which are populated with modern looking animal and plant fossils. Hominids of varying morphological similarity to homo sapiens are found throughout these layers, with fossils classified as archaic and then modern homo sapiens found in the most recent layers, dating as far back as several hundred thousand years... a fraction of a percent of the span represented by the column.
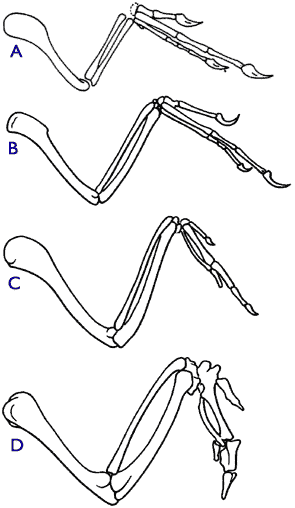
We never find mammal fossils embedded in pre-Carboniferous layers. We never find bird fossils in Permian layers. We never find primate fossils in Jurassic layers. We never find angiosperm fossils in the PreCambrian. We never find reptile fossils in the Ordovician layers. Etc.
This kind of distribution presents quite a distinctive pattern... which will be elaborated on as the discussion continues.


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}